
Usher Syndrome Part I, 2011 Edition: An Introduction to Sensory Perception
July 25, 2011
by Jennifer Phillips, Ph.D.
Note: this is the first of a four-part installment on the science of Usher syndrome. This series was posted on the Usher Syndrome Coalition Website and the ScienceBlogs network a few years ago, but given the amount of specific Usher science we discuss here on our blog, it may be helpful to readers to have these resources 'in house'. Part I gives an overview of hearing and vision at the cellular level, parts II and II discuss the known Usher genes and proteins, and their proposed roles in the eye and ear, respectively. Finally, Part IV brings all this cellular and molecular information back to the clinical level, and includes a survey of current and future research directions. Understanding how the human body works, and what goes wrong in any given disease, has value in and of itself. But for those of us hoping and striving for a treatment, a better understanding of the biology of Usher syndrome will be necessary for understanding how potential therapies might work.
Usher syndrome is a genetically recessive condition characterized by hearing impairment--usually from birth--due to defects in the sensory neurons of the inner ear, and vision loss due to retinal degeneration, which begins to occur in childhood or adolescence and progresses through several decades. Additionally, some Usher patients have balance problems associated with the sensory cell defects in the ear. There is a great deal of variation in the clinical presentation of the disease, and three clinical subtypes can be classified by the severity and age of onset of the symptoms. The latest estimates project that Usher syndrome affects about 1 in 6,000 people.
To begin to understand the pathology of this disease, one needs to focus on the affected cell types: Mechanosensory hair cells and photoreceptors. Both are highly specialized types of sensory cells, but they're performing essentially the same function, namely receiving an environmental stimulus and converting it into an electrical signal that is transmitted to the brain for interpretation. Although the nature of the stimuli-sound and light-are quite different, they are processed in much the same way, and thus it is not surprising to find a number of structural and functional similarities between photoreceptors and hair cells.
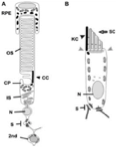
Figure 1: comparative anatomy of a photoreceptor cell (A) and a mechanosensory hair cell (B). The Outer Segment (OS) membranes of the photoreceptor are similar in form and function to the stereocilia (SC) of the hair cell. Both cell types contain cilia (labeled CC [connecting cilium] in A; KC [kinocilium] in B) and have specialized synapses (S) through which signals are sent to second-order neurons ('2nd', labeled in A; supporting cell projections can be seen at the bottom of the cell in B).
Sensory neurons are constantly stimulated with a complex array of information. Photoreceptors respond to all wavelengths of light within the visible spectrum as well as transmitting information about total light levels and movement. Mechanosensory hair cells can not only respond to physical contact by sound waves, they transmit information detailed enough to determine whether the sound waves in question were generated by a lover's whisper, breaking glass, or a bow being drawn across the strings of a cello. In order to intercept and convey information at this level of specificity, sensory cells have evolved specialized structures to meet the high demands of both input and output. On the receiving end are intricately organized membranes built to respond to the environmental signals. In photoreceptors, the outer segment contains stacked discs filled with opsin proteins, (remember those?) that trigger a cellular response when activated by light. In the hair cell, the sterocilia (not actually "cilia" at all, but finger- like projections made of the protein actin) move when they encounter sound waves, opening channels through which ions can enter the cell and initiate a response.
On the outbound side of things, these sensory neurons have a mechanism by which they can adequately relay the complexities of the environmental input they receive. In physiology 101, we learn about the classic type of neuronal response, in which the nerve cell needs to reach an action potentials to fire. Basically, these nerves are in a state of rest until they are sufficiently stimulated to elicit a response. Even though these responses to stimuli, such as a command from the brain in the case of motor neurons, or stepping on a sharp rock in the case of pain receptors, are fast, they are usually a simple binary, on/off type of response, and tend to be short lived.
The constant bombardment of information that our sensory cells endure is something along the lines of the chaos of the trading floor of the New York Stock Exchange, 24/7. A conventional neuron relying on action potentials to convey this information would be woefully overmatched in such a situation because it just wouldn't have time to 'reload' under such constant stimulation, nor would it be able to convey the specificity of information in the light or the sound stimulus receiving. Instead, photoreceptors and hair cells keep stockpiles of neurotransmitters tethered to the cell membrane. When the cell is stimulated by the environmental signal, no action potential needs to be achieved. Multiple neurotransmitters are right there, ready to be released, and, unlike a typical neuron, these sensory cells can be active in the long term-for as long as there is a stimulus to report. When these cells release neurotransmitters, they activate neurons waiting nearby to pass the message through additional channels leading to the sensory processing centers of the brain. The downstream message processing is far too complex to describe here, but the important thing to remember is that the message originates from these specialized sensory cells. They're responsible for collecting the information in the first place in a way that conveys a great deal of detail and specificity about the environment.
Finally, the presence of a true cilium is yet another commonality between these cells, although its function in each is quite distinct. So, in sum, we have two cell types that use similar cellular equipment to fulfill their roles as reporters of complex environmental information. We know that in cases of Usher syndrome, a genetic disorder, these two cell types are affected. This is strong evidence that the similarity between photoreceptors and hair cells goes deeper than the cellular similarities. In fact, the molecules regulating the specific sensory functions performed by these cells are yet another commonality, and this makes a lot of sense if you're trying to understand why both hearing AND vision are affected in Usher syndrome. It's all down to the molecular toolkits these cells use to develop and do their jobs properly.
In Part II, I'll introduce the proteins affected in Usher syndrome and describe what they tell us about the disease itself.






